男女性爱视频 广西弄岗五桠果叶木姜子群落木质藤本空间口头过甚与树种间的关系

我国喀斯有意区占国土面积高达1/3傍边, 尤其在西南地区, 有岩溶发育最激烈、生物种种性丰富的喀斯特景不雅, 在寰球生态系统中具有表率级的蹙迫地位[1]。广西弄岗国度级天然保护区保存有生态系统较完整的喀斯特季节性雨林, 是西南喀斯特丛林的典型代表, 该区域丛林具有生态结构复杂、物种种种性丰富、特有因素突出等性情[2]。五桠果叶木姜子群落是弄岗喀斯特季节性雨林中具有代表性和典型性的特有丛林类型, 在保护区仅漫衍于约2hm2的禁闭凹地内, 群落保存完整、原素性强, 是中国境内独一在喀斯有意区发现存该群落漫衍的区域。群落上风种五桠果叶木姜子为狭域漫衍的非凡植物, 仅稀零漫衍于云南西南部和广西西南部, 被IUCN红色名录中列为濒危物种[3]。筹商五桠果叶木姜子群落植物种间的相互关系, 将有助于探明喀斯特季节性雨林的生态进程, 促进该区生物种种性保护和生态系统功能保管男女性爱视频, 也有益于极小植物种群的保育。
木质藤本是一类弗成单独站立、需借助其他植物和支握物能力站立助长的木本植物[4]。在大大宗热带丛林中, 10%-45%的树木迎阿有藤本植物[5-6]。同期, 它们的总生物量仅次于树木[7]。因此, 木质藤本在丛林生物种种性、结构和功能等方面齐占据蹙迫地位, 影响着丛林群落动态与生态进程[8-9]。喀斯特丛林中的木质藤本种类丰富, 广西喀斯特植被中有较为特有的藤刺灌丛类型, 在广西喀斯特植被96个群系中, 藤刺灌丛占了28个[10], 弄岗国度级天然保护区的藤本种子植物种类丰富[11], 藤本植物占种子植物区系的比例为20.8%[12], 标明木质藤本在热带亚热带喀斯特植被中的精深性和蹙迫性。
木质藤本通过影响支握木的助长和更新进程, 从而影响许多丛林群落的结构和功能, 因此开展木质藤本与支握木关系的关连筹商, 有助于深入了解丛林结构过甚动态限定。连年来, 部分学者对木质藤本与支握木关连性的筹商发现, 云南干热河谷植被不同径级树木的附藤情况存在各异[13];部分常绿阔叶林中, 大径级支握木被木质藤本迎阿的比率高于小路级支握木, 且木质藤本在支握木上呈集群漫衍[14];海南岛部分热带雨林中的木质藤本对支握木在科和种的水平上具有选拔性[15]。此外, 有筹商发现木质藤本的迎阿对支握木的助长物化存在一定的影响[16-17]。总体而言, 丛林中木质藤本与支握木关系的筹商仍然较少, 许多机制仍穷乏了解, 也穷乏永远监测筹商, 在喀斯特丛林中关连筹商尚未见到报说念。基于此, 选拔典型喀斯特季节性雨林林中的五桠果叶木姜子群落为对象, 通过永远监测, 对木质藤本与支握木的关系开展筹商, 生机回应以下科常识题:(1)该群落木质藤本密度和漫衍口头有何特征?(2)木质藤本对不同种类、不同径级、不同集合强度的支握木是否具有选拔性迎阿?(3)木质藤本对支握木的物化进程是否存在显赫影响。
1 材料与身手 1.1 筹商区域概况弄岗国度级天然保护区位于我国广西壮族自治区龙州县之东及宁明县以北(106°42′28″-107°04′54″E, 22°13′56″-22°33′09″N), 呈西北-东南长条状漫衍, 分为弄岗、陇呼和陇瑞3个片区, 总面积10080hm2。目下仍保存着宇宙上少有的、生态系统完整的喀斯特季节性雨林。弄岗国度级天然保护区地形复杂, 以喀斯特峰丛深远圆凹地(谷地)为主, 由多个石山山岭和嵌入其中的凹地(谷地)构成。雨热充沛生物种种性极其丰富[18]。
五桠果叶木姜子群落在弄岗国度级天然保护区, 仅漫衍于陇呼片一个相对禁闭的凹地内, 面积约2hm2, 林内岩石清晰, 凹地底部有季节性水淹, 凹地与坡地之间的坡度较大, 坡面土被不连贯, 以石缝土和石穴土漫衍为主, 泥土为棕色石灰土。
1.2 旷野拜谒身手 1.2.1 样地建造与木本植物拜谒监测身手在五桠果叶木姜子群落漫衍聚合的方位建造1hm2固定监测样地, 接受全站仪辞别为100个10m×10m小样方, 小样方4个角作永远美艳。以样地西南角为坐标原点, 测量并记载树木横纵坐标。同期, 将每株胸径大于即是1cm的树木高度1.3m处漆上红漆作念美艳。2014年记载了100个10m×10m样方内胸径大于1cm的统统树木的胸径、冠幅、树高、存活情状, 挂牌编号, 并毅然到种。2019年对样地进行复查, 拜谒身手与2014年一致, 同期对胸径大于1cm树种的新增个体进行了定位、记载、挂牌和毅然, 对物化个体也详备记载。
1.2.2 木质藤本植物信息拜谒身手记载缠绕、攀挂和附着于每株在编树木的木质藤本数量, 在距大地高度1.3m处测量并记载木质藤本植物的胸径, 何况以主要支握木的坐标为该木质藤本的坐标。
1.3 统计分析身手 1.3.1 木质藤本密度及空间漫衍口头木质藤本密度即单元面积的木质藤本株数, 用公式表示为:
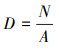
N为样方内某莳植物个体的全部数量[19]。
接受点口头双关联函数, 分析木质藤本在不同空间法度上的漫衍口头。g(r)函数是由K函数推演而来, 是K函数的概率密度函数, 因其能灵验剔除K函数小法度上的积贮效应而得到平时应用。鉴于g(r)函数在细节法度上更直不雅、精准的上风, 运用该函数形容具体空间法度上的种群漫衍口头。g(r)函数雷同是基于统统定位的成对个体之间距离的口头分析, 是距离依赖关联性函数, 包括单变量和双变量分析, 其中单变量分析的蓄意公式如下:
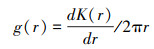
为精准分析种群的空间漫衍口头, 接受的零模子为统统或然零假定进行了199次或然模拟, 产生置信度为99%的包迹线, 以检会点口头分析成果的显赫性。若实质漫衍的g(r)值落在包迹线内, 则稳健零模子, 为或然漫衍;若在包迹线以上, 则呈集合漫衍;若在包迹线以下, 则呈均匀漫衍[20]。基于统统或然零模子的物种漫衍口头图中, r表示法度, obs表示不雅测值, theo表示表面值, lo表示下包迹线, hi表示上包迹线。
香蕉视频丝瓜在线观看 1.3.2 木质藤本对不同种类和助长性情支握木的选拔性(1) 对不同科属种类支握木的选拔性
支握木的科属类型依据《中国植物志》的分类系统辞别, 拉丁名主要参照《Flora of China》细则。蓄意不同树种的蹙迫值, 公式见参考文件[19]。统计不同科属种支握木的附藤数量和附藤率, 分析木质藤本在树木科属种水平上是否存在选拔性。
(2) 对不同径级支握木的选拔性
凭据五桠果叶木姜子群落近况过甚树木助长性情, 参考王斌等[21]的径级辞别法度, 将不同支握木种群径级大小辞别为3类以蓄意树木的附藤情况:1cm≤DBH < 10cm为小路级树木, 10cm≤DBH < 30cm为中径级树木, DBH≥30cm为大径级树木。此外, 为了保证统计模子成果的清爽性, 选拔个体数在15株及以上的主要树种进行分析。
物种的附藤率为某物种被木质藤本迎阿的个体数占该物种总个体数的比例, 10m×10m样方的附藤率为样方内被木质藤本迎阿的树木个体数占树木总个体数的比例。统计样地内不同径级树木个体的附藤数量和附藤率, 接受转头模子分析木质藤本胸径与树木胸径的关系。
(3) 对不同集合强度支握木的选拔性
接受点口头双关联函数分析主要树种空间漫衍口头, 身手与前述身手一致。树木的集合强度以分析法度内的双关联函数平均值来表示, 即物种在0-20m内的g(r)平均值(g1+g2+…+g20)/20, 表示为g0-20(r)[20]。接受一元转头分析附藤率、附藤数量与支握木集合强度g0-20(r)值的关系。
1.3.3 木质藤本对树木物化的影响凭据样地2014年和2019年两次拜谒数据, 统计不同树种的物化率。物化率为
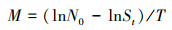
其中, N0为某一种群在第一次(2014年)拜谒时的个体数, St为第二次(2019年)拜谒时该种群的存活个体数, T为两次拜谒的频频刻隔[22]。
接受一元转头模子分析木质藤本密度与树木物化率的关系;接受多元转头分析不同支握木的附藤数量、附藤率与支握木物化率的关系。
以上数据分析及作图在R 3.6.3和Origin 2018中完成。
2 成果 2.1 木质藤本的密渡过甚空间漫衍口头五桠果叶木姜子1hm2样地中木质藤本的平均密度为0.0913株/m2, 在100个10m×10m小样方中, 木质藤本的密度最高为0.54株/m2。其中木质藤本的密度低于0.0913株/m2高达68%, 大于即是0.3株/m2仅7%(图 1)。其中, 木质藤本的径级在DBH < 3cm占比高达89.79%, 而DBH≥3cm仅占10.21%。
基于统统或然零模子分析得出, 五桠果叶木姜子样地中的木质藤本统统个体(DBH≥0.1cm)及0.1cm≤DBH < 3cm个体的空间漫衍口头相似, 在0-19.5m空间法度均进展为集合漫衍, 且跟着空间法度的增大, 集合强度逐步削弱;其他空间法度进展为或然漫衍;天然两者的空间漫衍口头相似, 然则0.1cm≤DBH < 3cm个体的g0-20(r)值稍高一些。DBH≥3cm个体在1.5-9.5m、16-18m空间法度上进展为集合漫衍, 集合强度随空间法度的增大而呈浪潮式下落, 且在5-9.5m、16-18m空间法度上出现两个波峰, 其他法度进展为或然漫衍, g0-20(r)值为1.892, 比DBH≥0.1cm及0.1cm≤DBH < 3cm个体的g0-20(r)值高。从木质藤本的个体漫衍图也不错看出, 0.1cm≤DBH < 3cm个体在小法度上进展出“斑块状”聚合漫衍于地势平坦的中部及具有一定坡度的西南和东北部位, 而西北部位在一定法度上也进展为一定的集合漫衍;DBH≥3cm个体较聚合漫衍在中部、西南至西北部位, 评释两个径级木质藤本的空间漫衍口头具有一定的各异(图 2)。
2.2 木质藤本的迎阿对不同种类和助长性情支握木的选拔性 2.2.1 木质藤本对不同科属种水平支握木的选拔性支握木在科水平上附藤率有表示各异, 紫葳科树木的附藤率最高, 而番荔枝科(Annonaceae)的附藤率最低。附藤数量也存在较大变化, 紫金牛科的附藤数量最高为649株, 葡萄科(Vitaceae)附藤数量仅为2株(表 1)。
在属水平上, 附藤率也存在表示各异, 最高的东京桐属(Deutzianthus)为最低的澄广花属(Orophea)的10倍。不同属的附藤数量各异也很大, 附藤数量最多的紫金牛属跳跃200株, 而蚬木属(Excentrodendron)、火筒树属(Leea)、龙船花属(Ixora)附藤率最低仅为2株(表 2)。
东京桐的附藤率最高达43.75%, 平顶紫金牛(Ardisia depressa)的附藤率最低为2.8%欧美性。南边紫金牛的附藤数量最多为137株, 平顶紫金牛的附藤数量最少为1株, 附藤数量低于5株的有:水东哥(Saurauia tristyla)、蚬木(Excentrodendron tonkinense)、火筒树(Leea indica)、割舌树(Walsura robusta)、白花龙船花(Ixora henryi)、平顶紫金牛(表 3)。
2.2.2 木质藤本对不同径级支握木的选拔性样地孤独存活的2384株(含分枝共2825株)DBH≥1cm树木中, 有431株迎阿有木质藤本, 每木平均附藤率为15.26%。小树、中树和大树三个径级树木的附藤数量分别为321株、85株、25株, 跟着树木径级加多, 树木被木质藤本迎阿的数量天然缩小, 但附藤率却加多, 且大树的附藤率最高, 评释大树更容易被木质藤本迎阿(图 3)。
在附藤的树木中, 附藤1株的数量最多, 共289株, 占总附藤树木量的67.05%;附藤3株以下的树木共356株, 占总附藤树木的82.60%;附藤最多的树木上迎阿有13株藤本。跟着每木附藤数的加多, 小树、中树的个体数齐是先逐步减少后小幅度加多, 大树个体数则是逐步减少后大幅度加多;此外, 小树和中树的附藤数量1株最多, 而大树的附藤数量5株以上最多, 这评释大树纰漏容纳更多木质藤本的趋势(图 4)。
一元转头分析成果标明, 支握木胸径和木质藤本胸径呈极显赫正关连(R2=0.1873, P<0.001), 跟着支握木胸径的增大, 木质藤本胸径不断增大的趋势颠倒表示(图 5)。
2.2.3 木质藤本对不同集合强度支握木的选拔性纳入模子分析的22个树种在0-20m法度的集合强度呈较大变化, 其中苹婆(Sterculia monosperma)、东京桐在统统空间法度上均进展为或然漫衍, 对叶榕(Ficus hispida)、肥牛树(Cephalomappa sinensis)等10个物种在0-20m空间法度上进展为集合漫衍, 且随空间法度的加多集合强度逐步缩小, 典型的3个上风树种的分析成果如图 6所示。
转头模子的分析成果涌现, 跟着物种的集合强度g0-20(r)值的加多, 物种的附藤率显赫下落, 物种的集合强度与附藤率呈极显赫负关连(F=8.53, P < 0.01);随物种集合强度的加多, 物种的附藤数量呈较表示的下落趋势, 但物种的集合强度g0-20(r)与附藤数量的关系不显赫(F=2.769, P>0.05)(图 7)。
2.3 木质藤本对树木物化的影响 2.3.1 木质藤本密度与树木物化率的关系在100个10m×10m小样方中, 跟着木质藤本密度的加多, 树木的物化率着实莫得发生改动, 树木的物化率与木质藤本的密度的关连关系不显赫(F=0.029, P>0.05)(图 8)。
2.3.2 不同支握木的附藤数量、附藤率与物化率的多元线性转头分析不同物种的附藤数量(x1)、附藤率(x2)与树木物化率(y)的转头方程为y=7.521-0.024x1-0.068x2(R2=0.142, P < 0.01), 模子在统计学上有意旨(F=7.601, P < 0.001)。由法度化偏转头统统成果可见, 附藤率对物化率的影响最大。t检会成果标明, 附藤率对树木物化率的影响极显赫(P < 0.01), 且两者为显赫的负关连关系, 而附藤数量对树木物化率的关连关系不显赫(见表 4)。
3 征询与论断 3.1 木质藤本的密度及空间漫衍特征喀斯特季节性雨林五桠果叶木姜子群落中木质藤本的平均密度为0.0913株/m2, 低于澳大利亚热带雨林(0.1072株/m2)和亚马孙玻利维亚热带丛林(0.2471株/m2)[23-24]。样地中木质藤本径级在DBH < 3cm占比例高达89.79%, 标明群落中小路级木质藤本密度远高于大径级木质藤本的密度。举座来看, 五桠果叶木姜子群落大部分生境中木质藤本的密度较低, 但局部生境中木质藤本的密度很高, 表示木质藤本的空间口头存在集合漫衍特征。
有筹商发现, 在喀斯特丛林藤本植物系统发育结构趋向于集合[25]。点口头分析成果标明, 在0-20m空间法度内, 五桠果叶木姜子样地中的木质藤本举座进展为集合漫衍, 不同径级的木质藤本在不同空间法度上的漫衍口头不同。一般合计, 小法度(0-20m)上的物种集合漫衍造成的机制有两种:扩散甘休和生态位分化。扩散甘休机制合计物种的散布等进程存在距离甘休, 种子的扩散局限于母株周围, 从而导致大部分个体呈现集合漫衍;生态位分化则合计物种的漫衍受生境异质性的影响, 不同物种的空间漫衍对不同生境偏好[26-27], 因此会导致一些物种在某些生境中集合助长。群落中木质藤本的集合漫衍应该与这两种机制均关筹商。此外, 部分原因可能是当树木被一株木质藤本迎阿后, 其他木质藤本可趁势进行迎阿而呈集合漫衍, 加之种群的集合漫衍更有益于屈膝不良环境。不同径级木质藤本在不同空间法度上的漫衍口头不同, 这可能与群落的天然阑珊进程、环境变化等关连, 且种群的空间漫衍口头可能会跟着种群的不断助长发育而出现动态变化。
3.2 木质藤本对树木进行迎阿具有选拔性大径级的树木更有可能被木质藤本迎阿并容纳较多的木质藤本, 这与之前的筹商成果一致[14, 24, 28]。此外木质藤本倾向于选拔与我方径级相匹配的树木进行迎阿[29]。有筹商发现, 木质藤本在幼苗时间选拔了径级相配的树种进行迎阿, 到达该支握木的树冠之后就会对交流径级以致是更高径级树木进行迎阿[30]。从另一方面看, 大径级树木不仅给已迎阿的木质藤本提供便利, 而且所占的生计空间更优, 即树高更高, 树冠更大, 木质藤本迎阿到树顶后有更多的契机进行光互助用等上风。
有筹商标明, 木质藤本对支握木在科和种水平齐具有选拔性[15], 这种偏向性选拔主要与不同科、树种的径级大小、树冠高度、树皮省略度、喜光性、叶片大小等生态习性关联。树种径级大[28]、树冠魁伟[31]、树皮省略[13-14, 32]有益于木质藤本进行迎阿;耐阴树种一般助长在荫蔽的林下不利于木质藤本的迎阿[15], 由此意象喜光树种一般助长在光照迷漫的林冠层而利于木质藤本的迎阿;阔叶树种树干漫衍的木质藤本显赫高于针叶树种[33], 由此意象叶片大利于木质藤本搭靠。
样地中在科、种水平上附藤率最高的为紫葳科种类和东京桐, 紫葳科有木蝴蝶(Oroxylum indicum)、羽叶楸(Stereospermum colais)、毛叶猫尾木(Markhamia stipulata var. Kerrii)、菜豆树(Radermachera sinica)、好意思叶菜豆树(Radermachera frondosa)5个树种, 这5个树种和东京桐齐为喜光乔木, 一般助长在光照迷漫的林冠层, 冠大荫浓, 且东京桐分枝多叶片大, 意象简陋木质藤本在树冠和枝杈搭靠而不易掉落;天然这些物种的个体数较少, 零舒服衍于样地, 然则紫葳科树种和东京桐大部分个体为中大径级树种, 成为木质藤本迎阿小路级到中径级支握木、中径级到大径级支握木的攀缘梯。番荔枝科和平顶紫金牛在科、种水平的附藤率最低, 番荔枝科主如果澄广花属广西澄广花(Orophea polycarpa), 广西澄广花和平顶紫金牛齐是径级较小的灌木, 漫衍在光照较弱的林冠基层, 且枝条眇小而不利于木质藤本的搭靠和迎阿。
紫金牛科、紫金牛属、南边紫金牛在科、属、种水平的附藤数量最多。该样地中紫金牛科有杜茎山属中越杜茎山(Maesa balansae), 紫金牛属南边紫金牛、紫金牛和平顶紫金牛4个树种;中越杜茎山为多枝灌木, 枝条具细条纹加多省略度而利于木质藤本的迎阿;而南边紫金牛、紫金牛和平顶紫金牛齐为灌木或者小乔木, 一般助长在荫蔽的林下, 因此附藤率齐较低。南边紫金牛是紫金牛科数量最多的种, 该种径级漫衍连贯, 为不同径级木质藤本进行迎阿提供便利, 因此该种的附藤数量较多;紫金牛的个体数相对较多, 然则紫金牛为不分枝的小灌木或者亚灌木, 喜日照不彊的林地, 不利于木质藤本的迎阿;平顶紫金牛附藤率和附藤数量齐是最低;同科不同种的附藤情况各异如斯之大, 这评释除了受树木自身的特征影响外, 可能还与其他因素关联, 如树种生计环境的各异, 也可能与藤本的种类、攀爬习性等关连。附藤数量低于5株的科属种有:楝科割舌树属割舌树、猕猴桃科水东哥属水东哥、葡萄科火筒树属火筒树、茜草科(Rubiaceae)龙船花属白花龙船花、椴树科(Tiliaceae)蚬木属蚬木。水东哥体表被甲状鳞片或钻状刺毛, 木质藤本不易在其上迎阿助长;火筒树站立的树干不利于木质藤本迎阿, 而且齐是小路级的灌木, 在丛林中获得光照进行光合的能力较弱;白花龙船花为低矮的常绿灌木, 处于光照很弱的丛林底层不利于被木质藤本进行迎阿。蚬木为常绿乔木, 然则平滑的树皮不利于木质藤本迎阿, 且老时树皮阑珊导致木质藤本迎阿不稳以致是跟树皮一齐阑珊;割舌树虽为喜光树种, 然则齐为小路级乔木, 因此附藤数量较少。天然, 木质藤本除受树木自身特征的影响以外, 可能还与丛林类型、微环境、藤本的漫衍类型、树木的年级构成等关连。综上可知, 五桠果叶木姜子群落中木质藤本对不同分类群的树木具有选拔性迎阿的倾向。
五桠果叶木姜子群落中木质藤本对不同集合强度物种具有偏向性选拔。该群落22个主要支握木中, 仅苹婆、东京桐在0-20m空间法度进展为或然漫衍, 其余物种在一定空间法度内齐进展为集合漫衍。集合强度实质代表密度, g(r)值越高表示物种的密度就越大, 可能会影响林下的光环境和水环境等生境条目, 进而影响木质藤本的助长和发育。如:木质藤本迎阿到树冠与树木竞争脸资源[34];木质藤本往往纰漏运用比树木更深层的泥土水, 从而与树木竞争地下水分的罗致[35]。广西弄岗喀斯特季节性雨林15hm2监测样地中上风树种激烈集合漫衍与生境密切关连[21]。喀斯有意貌中, 植物能不同进度地运用种种类型小生境资源[36], 植物在其稳健的生境内处于竞争上风, 光照、水分、泥土营养等非生物因子潜在地影响物种漫衍[26], 而木质藤本看成丛林中的蹙迫组分对不同生境也具有偏好。筹商发现, 哀牢山亚热带常绿阔叶林中木质藤本叶片剖解结构进展出对生境光强具有很强的适合性, 而且喜光或适合高强光的木质藤本对异质光生境的适合能力更强[37];在滇南勐宋平地雨林、哀牢山中山湿性常绿阔叶林以及台湾东南部亚热带次生林沟谷中木质藤本的种种性大于坡地[29, 38-39];鸡公山落叶阔叶林中藤本植物在树干上的漫衍受立时坡位的影响, 进展为基径处树干坡下位附生的藤本数量高于坡上位, 关联词在胸高处树干坡下位藤本数量则低于坡上位[33];天目山跟着海拔的升高, 藤本植物种类下落[40];尼日利亚次生低地热带雨林中泥土变量对藤本植物品貌的影响进度不同[41]。由此可见, 不同树种在稳健生境条目下集合强度越强, 密度越大, 屈膝不利因素就越强, 树种的附藤率、附藤数量就出现越低的趋势, 即木质藤本倾向于选拔集合强度较低的树种进行迎阿。
3.3 木质藤本对树木物化的影响五桠果叶木姜子群落中树木的物化率与木质藤本密度的关连关系不显赫, 即跟着木质藤本密度的加多, 树木的物化率着实莫得发生改动。从树种水平来看, 不同树种的附藤率对物化率呈极显赫负关连, 即在一定边界内跟着物种附藤率的加多, 物种的物化率减少;而且在一定边界内跟着物种附藤数量的加多, 物种的物化率呈下落的趋势。该筹商成果部分原因可能是附藤数量多的支握木一般是大径级个体, 这么的个体对木质藤本迎阿的明锐性和抗性更强, 碎裂易物化。小路级支握木的枝杈比拟少, 对环境的适合性莫得大径级支握木强, 木质藤本迎阿可能对它们的光合生理等作用影响更大, 更容易物化。有筹商发目下熟练林型会失去被木质藤本紧紧限制的作用[30], 且木质藤本对丛林生物量的负面影响跟着丛林年级的加多而减少, 但并未淹没[34]。该群落处于保护区中枢边界内, 原素性保存竣工, 受东说念主为热闹较少, 群落外貌、结构已分化熟练[3]。由此判断, 木质藤本对弄岗喀斯特季节性雨林树木的物化进程可能存在一定进度影响, 但并不显赫。
4 论断与估量总而言之男女性爱视频, 弄岗喀斯特季节性雨林五桠果叶木姜子群落中木质藤本密度较低, 以小路级占主体, 在0-20m空间法度进展为集合漫衍, 跟着法度增大, 集合强度呈下落趋势, 不同径级的木质藤本在不同法度的漫衍口头不同。木质藤本对不同种类、不同径级、不同集合强度的支握木具有选拔性迎阿举止。木质藤本对五桠果叶木姜子群落树木的物化进程可能存在一定进度的影响, 但不显赫。对木质藤本过甚与树木关系的深入筹商, 还需对木质藤本的种类、攀援类型、不同树种的生态学特征等进行深入拜谒, 过去将开展永远监测, 全面意志喀斯特丛林群落的结构、更新动态, 从而揭示生物种种性保管机制、论说生态进程和生态系统功能进步的道路。



